

Article written by: Jayson Kowinsky - Fossilguy.com
Allosaurus
Fast Facts about Allosaurus

Rendering of Allosaurus fragilis by Fred Wierum (CC BY-SA 4.0)
Name:
Allosaurus (pronunciation: "Allo-SAWR-us")
The name means "Different Lizard" - due to the different vertebra compared to other dinosaurs discovered at the time.
Taxonomy:
Dinosauria (Dinosaur) - Saurischia (Lizard Hipped) - Theropoda (Beast Footed)- Allosauridae (Family) - Allosaurus (Genus) - A. fragilis, A. jimmadseni, A. europaeus (species)
Age: Late Jurassic
Allosaurus is found in the Morrison formation (155-148 myo) of North America and the equivalent Lourinha formation (152-146 myo) of Portugal.
Extinction: Late Jurassic
Allosaurus went extinct with many other dinosaurs during the faunal turnover of the Late Jurassic.
Discovery: Ferdinand Hayden, 1869
A broken tail vertebra of a possible Allosaurus was given to geologist Ferdinand Hayden during his geological survey of Colorado.
Distribution: North America and the Iberian Peninsula (Southwest Europe)
Allosaurus had a wide geographic range across North America and also into Western Europe.
Body Size: Large: over 40 feet (13 meters)
It was one of the largest Theropods of the Jurassic. The average length was less than 30 feet in length and Madsen (1976) estimates the largest known specimen (AMNH 5767) to be 13m (42.6 feet).
Diet: Carnivore
Allosaurus was an apex predator. It hunted other large dinosaurs including Sauropods and Stegosaurus.
Speed: Fast: 21 mph (9.4 m/s)
Allosaurus had long muscular legs and was faster than other dinosaurs of the time, including the Sauropods that it hunted.
A 2007 study by Sellers et al. gives a maximum speed estimate of 9.4 m/s (21 mph).
Fun Facts:
Allosaurus is the most abundant large theropod in the Morrison Formation
Allosaurus' jaws were incredibly weak, it had unusual neck muscles, and could open its mouth at a very wide angle.
This evidence suggests it ate like a bird of prey, by grasping and tugging flesh from the animal.

Overview of Allosaurus

An Allosaurus in pursuit of a Stegosaurus. Display at the Los Angeles Natural History Museum.
Allosaurus was one of the largest and most abundant theropods of the Late Jurassic in North America. It has a wide geographical range and is found throughout the Morrison Formation which was deposited around 155 to 148 million years ago. It also crossed into Europe and is found in the Lourinha formation of the Iberian Peninsula (Portugal and Spain) which has a similar dinosaur fauna to the Morrison.
Allosaurus was an apex predator. Although the average size was just shy of 30 feet, the largest known specimen, AMNH 5767, has an estimated length of 13m or 42.6 feet (Madsen 1976). Allosaurus is characterized by a large and slender head, crests over its forward-facing eyes, and many dagger-like serrated teeth. This means it had good depth perception for hunting prey and could easily tear into flesh. It has also been found to have a large olfactory bulb, which indicates a good sense of smell. Allosaurus has relatively long arms with 3 large claws on each hand. This allowed it to grasp onto and tear into struggling prey. It also has a long tail and muscular legs that are built for speed. Estimates show it could run up to 21 miles per hour (Sellers et al. 2007), which was faster than other dinosaurs of its time, including the Sauropods and Stegosaurs it hunted.
We know it preyed on many of the larger dinosaurs of the Morrison formation such as Stegosaurs and Sauropods based on feeding evidence and injuries. There have been Allosaurus bite marks found on Stegosaurus back plates and Apatosaurus vertebra. There have also been injuries found on Allosaurus bones made from the tail spike of a Stegosaurus (Bakker et al. 2014). Evidence from the Mygatt-Moore Quarry even shows signs of cannibalism from scavenging on other Allosaurus in a stressed environment (Drumheller et al. 2020).
A surprising fact about Allosaurus is that it had very weak jaws. It was not built for crushing bone like some other large theropod dinosaurs. Its slender jaws had a weaker bite force than that of a crocodile and a lion. However, Allosaurus had a long and mobile neck and it could open its jaws over 90 degrees! This evidence suggests that it ate, not by biting and ripping off chunks of meat and bone, but instead by striking and picking at it like a falcon (Snively et al. 2013). It could grab flesh, then quickly and forcefully pull its head back to de-flesh the prey.
There were at least 3 species of Allosaurus at various times in the late Jurassic. By the end of the late Jurassic, Allosaurus went extinct with many of the other dinosaurs during the faunal turnover leading into the Cretaceous.

A fossil pubis bone of an Allosaurus showing an entry and exit wound from a stegosaurus tail spike. The white plaster is a cast of the abscess formed from infection after the injury. Image Credit: Robert Bakker
The Jaws: A Unique Feeding Style:
Not a hatchet attack, but a falcon-like strike-and-tear model

Allosaurus skull: Showing the maximum possible gape of Allosaurus specimen AMNH666 as hypothesized by Robert Bakker. Image by: Steveoc 86(CC BY-SA 3.0)
Since Allosaurus was a large apex predator, many people think it had powerful bone-crushing jaws similar to the Tyrannosaurs of the Cretaceous. However, its skull, jaws, neck, and teeth, were uniquely designed for a very different feeding strategy.
In 2001 Rayfield et al. modeled the bite force of Allosaurus and found it to be a mere 3,572 N (803 lbs). This is very small. For example, a lion, which is tiny in comparison, has a larger bite force at 4,167 N (936 lbs). Tyrannosaurus rex has an astounding bite force of 13,400 N (3,012 lbs). Rayfield et al. found that although the jaws were very weak, the narrow cranium could withstand high forces, which suggests a "very specific feeding behavior."
What is this specific feeding behavior?
One feeding strategy proposed was a "hatchet attack" where Allosaurus would open its jaws very wide and swing downward with its upper jaws, like a hatchet, causing massive damage for each blow. This attack style has become particularly popular after Stephan Lautenshlager used software to model musculoskeletal structures and muscle strain around the jaws to determine the jaw gape of different theropods (2015). He found that Allosaurus had a maximum jaw gape angle of 92 degrees, which is incredibly wide. An animated model of this gape is shown in the Sketchfab below. It's important to note, however, that the 92 degree angle is the maximum gape, and the optimal gape was found to be a mere 32.5 degrees.
Unfortunately, people tend to ignore the optimal gape and concentrate on the maximum gape, and then use that to justify a gruesome hatchet style attack. In reality, the hatchet style attack doesn't work. Anton et al. (2003) tested numerous attack styles based on Allosaurus' unique skull, neck, and tooth characteristics. They found a hatchet attack to be implausible for many reasons. One of the reasons is Allosaurus has relatively small teeth compared to similar sized theropods, and a significant number of them would have broken during every "hatchet" type attack. The dinosaur would not have been able to replace them fast enough and would have been toothless.
The attack model that Anton et al. (2003) found to work the best was a strike-and-tear model that birds of prey use today. They found the low crowned teeth would allow it to grab the prey without the need for a large gape. The smaller jaw bite force would be compensated by the contribution of the unique neck muscles that would help push the jaws into and out of the prey. Anton et al. also adds the powerful arms and large claws would have helped hold onto the prey to provide stability and control during biting. Another interesting finding was the strike-and-tear model worked best on slightly convex surfaces, meaning very large prey animals, like Sauropods and Stegosaurs (which we know it ate).
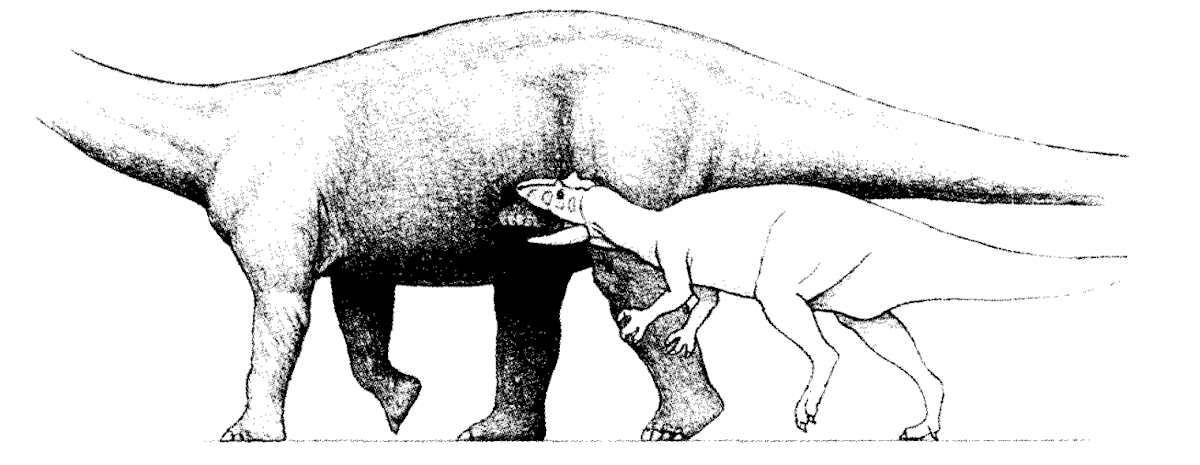
Figure 5 from Anton et al. 2003 showing the most plausible feeding strategy. The bite is powered both by the jaw and the neck muscles.
Snivley et al. (2013) also looked at Allosaurus' specific feeding behavior. They made really cool multibody dynamics models of Allosaurus and also came to a falcon-like strike-and-tear feeding model. Finally, Lautenshaget (2015) found the wide jaw gape simply helped it reduce the risk of jaw dislocation, which is important for a strike-and-tear mode to attack prey. The Allosaurus strike-and-tear mode is very similar to what modern falcons do today.
For your viewing pleasure is an animated 3D sketchfab of its gape, and a video showing its feeding strategy:
3D INTERACTIVE MODEL - Animated model of an Allosaurus fragilis skull showing the extent of it's gape. - Tap to Start Model
Interactive 3D Model of Allosaurus fragilis by Thomas Flynn (CC0 Public Domain) on Sketchfab. This model uses Stephan Lautenschlager's allosaurus gape data from his 2015 paper.
YOUTUBE VIDEO: Allosaurus Feeding Movements
Animation of the feeding movements of the head and neck of the Jurassic predatory theropod dinosaur Allosaurus based on multibody dynamics and soft-tissue reconstruction. Based on Snivley et al., 2013. By: WitmerLab at Ohio University
Discovery and Naming of Allosaurus
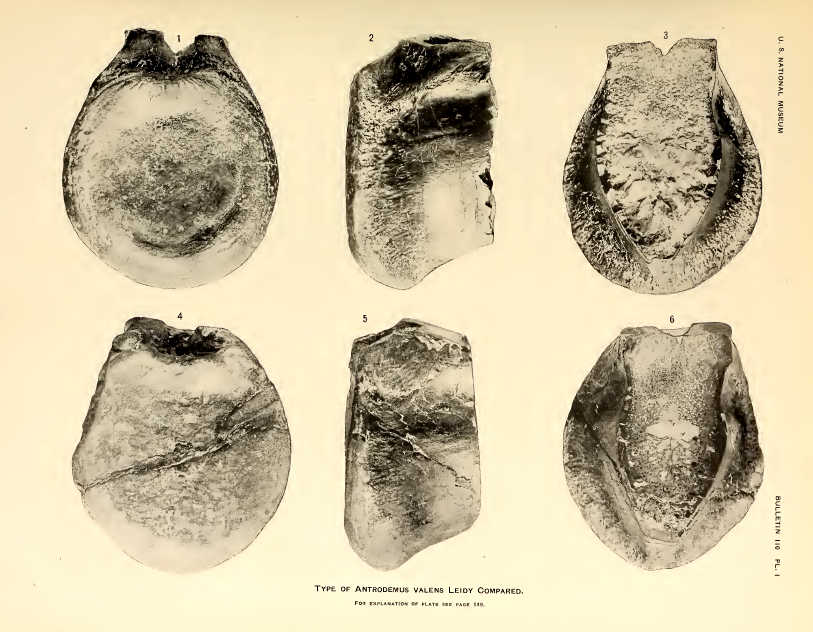
The type specimen of Antrodemus that Leidy described in 1870. From Plate 1 of USNM Bulletin 110, 1920 (Public Domain)
The discovery of Allosaurus starts in 1869 when local residents of Middle Park, Colorado gave a small, broken tail vertebra to geologist Ferdinand Hayden. This vertebra made its way to Joseph Leidy who described it in 1870. Leidy named it Poekilopleuron, after a known European dinosaur. A few years later in 1873, Leidy realized it might be different and changed its name to Antrodemus. A problem with this specimen was the exact location was never specified, so it could have come from surrounding Cretaceous rock, meaning it might actually be a T. rex vertebra.
Many more Allosaurus discoveries were made during the Bone Wars of the 1870's. This is where Cope and Marsh frantically created new dinosaur names out of every little bone they could find. Out of the craziness came many names for this Jurassic theropod including Allosaurus fragilis, Epanterias amplexus, Allosaurus lucaris, Hypsirophis discursis, Creosaurus atrox, and Labrosaurus sulcatus.
Overall, the original name Antrodemus stuck, and it wasn't until 1976 that Allosaurus fragilis became the official name. In 1976 James Maddison published a paper saying the original Antrodemus fossil was too fragmentary and did not have sufficient location data for it to be an official name. He chose Allosaurus fragilis based on better fossil material that Marsh had named in 1877.
Allosaurus Species
Today, 3 species of Allosaurus are generally accepted: A. fragilis, A. jimmadseni, and possibly A. europaeus. In the past, there were as many as 19 named species. However, studies such as the one from Carpenter (2011), show there is tremendous variation between individual allosaurus specimens. Variations can be caused by age, sex, population, and simply unique individual differences. As a result, all of the species except for the three are generally deemed invalid.
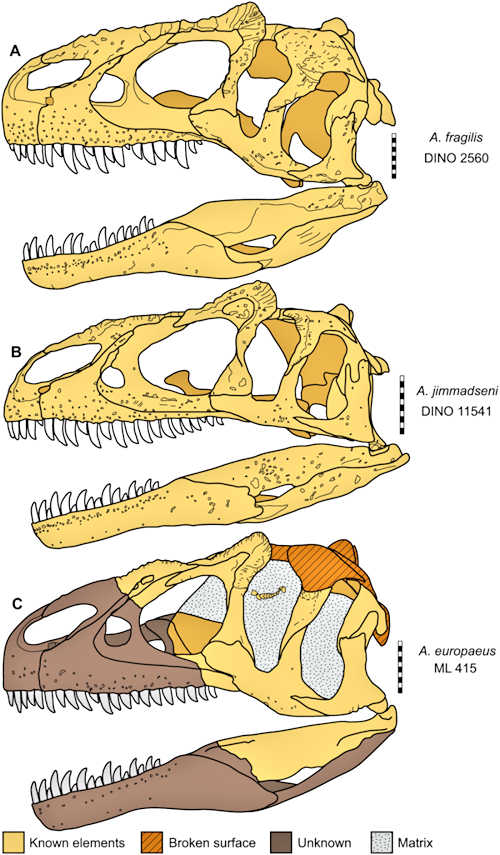
A comparison of the skulls of the 3 Allosaurus specimens. The caption reads: (A) Allosaurus fragilis (DINO 2560). (B) Allosaurus jimmadseni (DINO 11541). (C) Allosaurus europeaus (ML 415). Scale bars equal 10 cm. This is Figure 16 from Chure and Loewen 2020 (CC BY 4.0)
Allosaurus fragilis
A. Fragilis, found throughout the Brushy Basin Member of the Morrison Formation, is by far the most common allosaurus. This is the younger species found in the Morrison Formation. Although the species are very similar, this species is better suited for hunting and tackling large prey than the older species. The skull is more robust and has better binocular vision, which is crucial for hunting prey.
Allosaurus jimmadseni
A. jimmadseni is an older species found in the Salt Wash Member of the Morrison Formation. This species is less common and occurs a few million years earlier than A. fragilis. It was named in a paper by Chure and Loewen in 2020. The species name refers to James Madsen, the Paleontologist who studied and excavated 1000's of Allosaurus bones from the Cleveland-Lloyd Dinosaur Quarry in Utah.
The holotype of A. jimmadseni is called DINO 11541 and was found at Dinosaur National Monument in Utah. Two other notable specimens are 'Big Al' (MOR 693), and 'Big Al II' (SMA 0005) both of which were found in or adjacent to the Howe Quarry in Wyoming.
Chure and Loewen (2020) go over a number of differences between A. jummadseni and A. fragilis. Overall, A. jummadseni has a less robust skull and is less well suited for tackling larger prey. One main skull difference from A. fragilis is that it has a straighter jugal bone (the cheekbone). It also has differently shaped skull openings, including the eye orbitals and the fenestrae. This shows the binocular vision was not as good as in A. fragilis. The bones on the rear of the skull are connected differently between the two species. A. jummadseni also has more crests. The crest near the eye is larger and it also has pronounced blade-like crests on its nasal area.
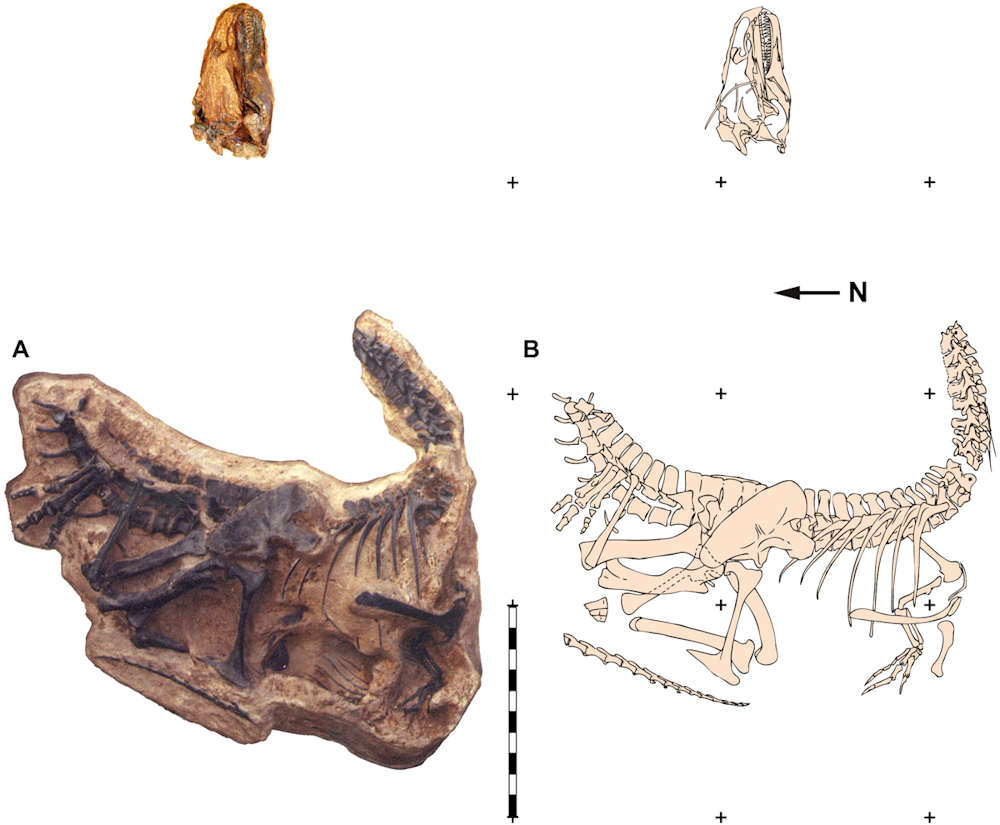
The holotype specimen of Allosaurus jimmadseni (DINO 11541). Photos by Dan Chure. Scale bar equals one meter. This is Figure 1 from Chure and Loewen 2020 (CC BY 4.0)
Allosaurus europaeus
A. Europaeus is found in the late Jurassic Lourinha Formation of Portugal. During the late Jurassic, the Atlantic Ocean was new and very narrow. North America and the Iberian peninsula (Spain and Portugal) were especially close together. Dinosaurs could easily have crossed between the continents. As a result, the dinosaur fauna of the Lourinha formation is very similar to the dinosaurs of the North American Morrison formation.
Fossils of an Allosaurus were originally discovered near Valada, Portugal in 1999. The fossils consisted of a partial body that was missing the head. The initial identification was Allosaurus fragilis. A few years later, a partial skull (ML415) was found along the coast near Lourinha. The skull was found in the Porto Novo Member of the Lourinha Formation, which is almost the same age as the Brushy Basin Member of the Morrison Formation. This skull was studied and a paper was published in 2006 by Mateus et al. showing the skull had characteristics different from A. fragilis, and they named the specimen A. europaeus, which means the European Allosaurus.
Not everyone agrees that A. Europaeus is a valid species. Evans et al. (2020) examined the holotype of A. europaeus and compared it to numerous specimens of A. fragilis. They found the unique diagnostic characteristics of A. europaeus were also present in some A. fragilis specimens. Meaning, A. europaeus could simply be A. fragilis, and not a different species. It looks like a better skull is needed to determine if this is indeed a new species, or simply A. fragilis.
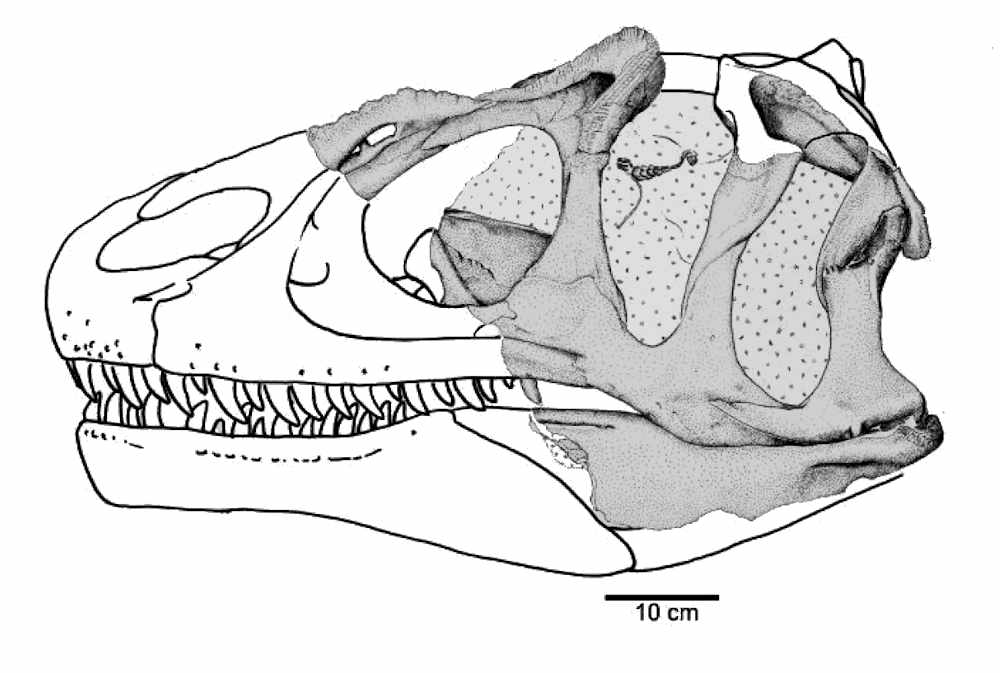
The holotype specimen of Allosaurus europaues (ML415). The shaded regions are the fossil parts that were found. This is Figure 7 from Mateus et al. 2006
Notable Quarries and Specimens
The Morrison formation had a seasonably arid climate with widespread droughts, making it a harsh environment. During droughts, some sauropods may have migrated, but others would have congregated near water sources, which helped create numerous dinosaur bone beds that dot the Morrison formation. The quarries below highlight some of the more famous bone beds with notable Allosaurus fossils.
Cleveland-lloyd Dinosaur Quarry:
Largest concentration of Allosaurus bones

Picture of Dinosaur bones (mostly Allosaurus) at the Cleveland-Lloyd Dinosaur Quarry. Image Credit: gjhikes.com
The Cleveland-Lloyd Dinosaur Quarry in central Utah contains the densest concentration of Jurassic dinosaur bones yet discovered. Many of the 1000's of bones there are from Allosaurus, and at least 46 individuals have been discovered in the quarry (Carpenter 2010). This is highly unusual, as the most common bones in the Morrison formation are Sauropods, not carnivorous theropods.
Many theories have been proposed as to why are there so many predatory theropods in one place. One of the original theories was that it was a classic predator death trap, where an animal would get stuck in the shallow muddy lake, and Allosaurus dinosaurs would attempt to go in and eat the animal, becoming trapped themselves (sort of like the La Brea Tar Pits). However, a newer theory suggests something else.
In 2017 Peterson et al. put forth a new model on how so many Allosaurus bones ended up in one place. They noted there was a lack of typical aquatic fish and reptiles, indicating the water was toxic, possibly due to rotting dinosaur carcasses.
The bones were mostly disarticulated and many did not show signs of predation. There is also a lack of shed Allosaurus teeth, which would indicate a feeding ground. Many of the bones also showed signs of weathering. All of this pointed to some type of flood that washed the carcasses to this area. The rotting carcasses would have made the water toxic for aquatic animals. The dinosaurs rotted and the bones accumulated. The orientation of the bones also show at least 3 separate flood events that washed animals into this place, indicating seasonal deposition that built up the massive bone bed. This doesn't quite explain everything, but it's the best theory so far.
The Howe Quarry:
Home of Big Al and Big Al II
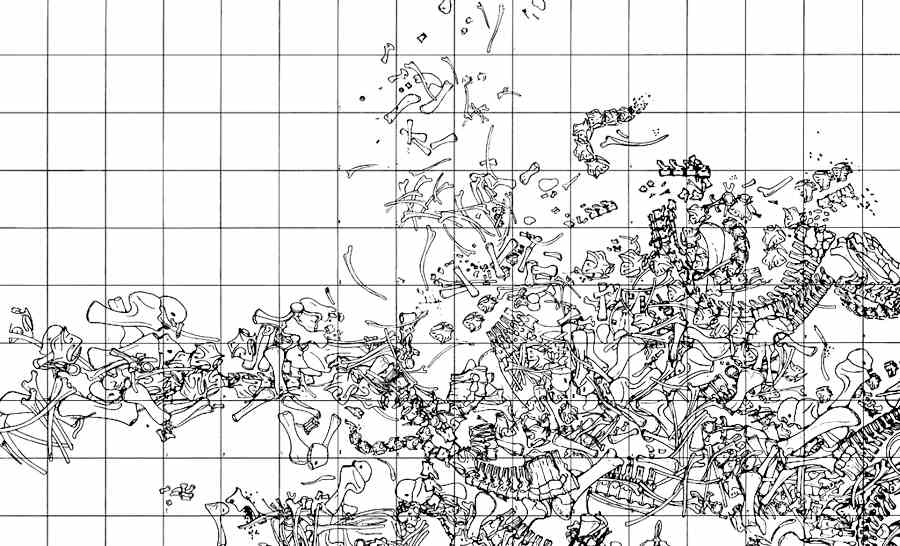
This is a historical map of some of the dinosaur bones excavated by Barnum Brown in the Howe Quarry: Circa 1930 (Public Domain)
In 1934 Barnum Brown (Discoverer of T. rex) found this incredibly rich dinosaur graveyard. The original excavation was only 45 x 65 feet across, but It contained over 4000 dinosaur bones. Many of them appear to be massive Sauropods that got stuck in mud during a drought. There are also numerous shed Allosaurus teeth, indicating they fed on the Sauropods, and a few Allosaurus skeletons.
Two remarkable Allosaurus jimmadseni specimens found here are Big Al (MOR 693) and Big Al II (SMA 0005). In 1991 Big Al was found accidentally on BLM land just outside of the Howe Quarry by an international research team. There they found a 95% complete Allosaurus skeleton which measured 26 feet in length. Big Al had numerous broken bones and bone infections; he was in bad shape when he died.
In 1996 the same team came back to the quarry to find another Allosaurus, this time in the actual quarry. Here they found another large and even more complete specimen they named Big Al II. Together, these are the two most complete A. jimmadseni specimens to date.

This is a cast of "Big Al" on display at the Museum of the Rockies. Image by: Tim Evanson (CC BY-SA 2.0)
Recommended Dinosaur Books and Educational Items:

The Rise and Fall of the Dinosaurs: A New History of a Lost World
Steve Brusatte, 2019
This dinosaur book is light-hearted and easy to read. The author, a renowned Paleontologist, does a great job at engaging the reader. It is packed with details without being overwhelming. Check out the reviews; it's a great new dinosaur book! Available in Kindle, Paper, and Audio.


Dinosaurs Rediscovered: The Scientific Revolution in Paleontology
Michael J. Benton, 2019
This is a great general book on dinosaurs. It's up-to-date and goes over the new discoveries over the last 20 years that has completely changed our views on dinosaurs. The book is broken up into easy chapters that goes over their colors, speeds, bite force, child caring, and more... stuff we didn't know just years ago! Check out the reviews and the sample pages. Available in Kindle, Paper, and Audio.

Nifeliz Dinosaur Fossils Building Kit
From 8 dinosaurs to choose from, The Nifeliz Dinosaur sets are nearly 500 pieces. They are a great gift for those 12+ who enjoy solo or group building with LEGOs. Constructing these models enhances hand-eye coordination and creativity. Perfect for family or friends, It's also compatible with existing LEGO pieces, facilitating the creation of a personalized toy museum! Check out the reviews!

High quality Dinosaur teeth by Fossilera
 Paleoadventures Dinosaur Digs
Paleoadventures Dinosaur Digs
References / Works Cited
Anton, M., Sanchez, I.M., Salesa, M.J., & Turner, A. (2003) The muscle-powered bite of allosaurus (dinosauria; theropoda): an interpretation of cranio-dental morphology. Estudios Geologicos-madrid, 59, 313-323. http://dx.doi.org/10.3989/egeol.03595-6106
Bakker, Robert T.1, Zoehfeld, K. Weidner1 and Mossbrucker, Matthew T. (2014) Stegosaurian martial arts: a jurassic carnivore stabbed by a tail spike, evidence for dynamic interactions between a live herbivore and a live predator. 2014 GSA Annual Meeting in Vancouver, British Columbia (19-22 October 2014) Paper No. 221-6
Bates, Karl & Falkingham, Peter (2012). Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics. Biology letters. 8. 660-4. doi.org/10.1098/rsbl.2012.0056
Carpenter, Kenneth (2011) Variation in a Population of Theropoda (Dinosauria): Allosaurus from the Cleveland-Lloyd Quarry (Upper Jurassic), Utah, USA. Paleontological Research. 14. 250-259. doi.org/10.2517/1342-8144-14.4.250
Chure DJ, Loewen MA. (2020) Cranial anatomy of Allosaurus jimmadseni, a new species from the lower part of the Morrison Formation (Upper Jurassic) of Western North America. PeerJ 8:e7803 doi.org/10.7717/peerj.7803
Drumheller SK, McHugh JB, Kane M, Riedel A, D'Amore DC (2020) High frequencies of theropod bite marks provide evidence for feeding, scavenging, and possible cannibalism in a stressed Late Jurassic ecosystem. PLOS ONE 15(5): e0233115. doi.org/10.1371/journal.pone.0233115
Evers, S. W., Foth, C., & Rauhut, O. (2020) Notes on the cheek region of the Late Jurassic theropod dinosaur Allosaurus. PeerJ, 8:e8493. doi.org/10.7717/peerj.8493
Lautenschlager Stephan (2015) Estimating cranial musculoskeletal constraints in theropod dinosaurs. R. Soc. open sci.2150495
doi.org/10.1098/rsos.150495
Madsen, James H., Jr. (1993) [1976] Allosaurus fragilis: A Revised Osteology, Utah Geological Survey Bulletin 109 (2nd ed.), Salt Lake City: Utah Geological Survey from Utah Dig. Library
Mateus, Octavio & Walen, Aart (2006) The large theropod fauna of the Lourinha Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus. New Mexico Museum of Natural History and Science. Bulletin 36:123-12936. Article Here
Rayfield, E. J., Norman, D. B., Horner, C. C., Horner, J. R., Smith, P. M., Thomason, J. J., and Upchurch, P. (2001). 'Cranial design and function in a large theropod dinosaur', Nature, 409, pp. 1033 - 1037. Accessed 24th August 2019. doi.org/10.1038/35059070
Sellers William Irvin and Manning Phillip Lars (2007) Estimating dinosaur maximum running speeds using evolutionary robotics. Proc. R. Soc. B.2742711-2716. doi.org/10.1098/rspb.2007.0846
Snively, Eric, Cotton, John R., Ridgely, Ryan, and Witmer, Lawrence M. (2013) Multibody dynamics model of head and neck function in
Allosaurus (Dinosauria, Theropoda), Palaeontologia Electronica Vol. 16, Issue 2; 11A 29p;
palaeo-electronica.org/content/2013/389-allosaurus-feeding doi.org/10.26879/338
Storrs, G., Oser, S., & Aull, M. (2012) Further analysis of a Late Jurassic dinosaur bone-bed from the Morrison Formation of Montana, USA, with a computed three-dimensional reconstruction. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 103(3-4), 443-458. doi.org/10.1017/S1755691013000248






